- circRNA Molecules Have, Oddly, No Beginning or End
- circRNA Are Now Recognized as Regulators of Gene Expression
- A Flurry of New Findings Indicate circRNA Are Also Templates for Synthesis of Proteins Having As Yet Unknown Functions
About a year ago, my blog titled Curiously Circular RNA pointed out that circular RNA (circRNA) in animals are odd molecules in that, unlike the vast majority of other RNA in animals, circRNA have no structural beginning (5’) or end (3’). This very curious feature has, not surprisingly, stimulated considerable scientific interest in knowing more about these molecules, which were serendipitously discovered some 30 years ago.
Application of next-generation sequencing has revealed that circRNA are actually relatively abundant and evolutionarily conserved, which implicates biological importance rather than inconsequential mistakes during RNA splicing mechanisms. Some circRNA have been shown to have function—circRNA can hybridize to complementary microRNA (miRNA), and thus serve as a kind of ‘sponge’ that influences miRNA-based gene expression. Evidence for circRNA involvement in gene expression continues to grow, as there are now >700 items on “circRNA [and>
sponges” in Google Scholar.
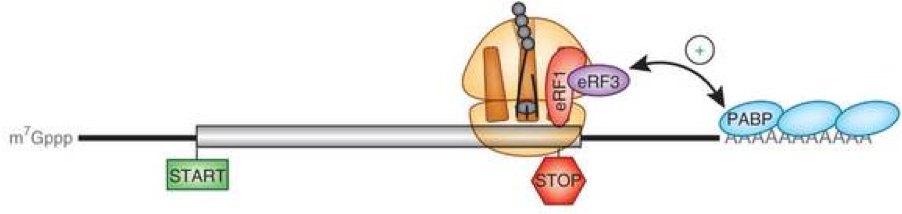
Very recently published lines of research (that I’ll outline in what follows) implicate circRNA as coding templates for proteins, which heretofore has been exclusively associated with messenger RNA (mRNA). Current dogma holds that translation of mRNA into protein requires recognition of the 7-methylguanylated (m7G) 5’-cap structure to start ribosome binding, while the 3’-poly(A) tail protects the mRNA molecule from enzymatic degradation and aids in stopping translation, as depicted below.
Start and stop structural elements characteristic of mRNA are obviously not present in circRNA, which are literally just circles of RNA. Consequently, finding proteins encoded by circRNA has stirred up controversy about whether such proteins are a new and fundamentally important aspect of genetics or just inconsequential biochemical mistakes.
Translation of circRNA in Fly Head Neurons
 Fruit fly. Taken from turbosquid.com
Fruit fly. Taken from turbosquid.com
Researchers at The Hebrew University of Jerusalem in Israel in collaboration with a team at Max-Delbruck-Center for Molecular Medicine in Berlin, Germany recently reported in Molecular Cell the first compelling evidence that a subset of circRNA is translated in vivo. The study by Kadener & coworkers was carried out using the common fruit fly (Drosophila melanogaster), which is known to have a number of features that lend to investigations of circRNA: (1) >2,500 fruit fly circular RNAs have been rigorously annotated, (2) these are mostly derive from back-splicing (pictured below) of protein-coding genes, (3) hundreds of which are conserved across multiple Drosophila species, and (4) exhibit commonalities to mammalian circRNA.
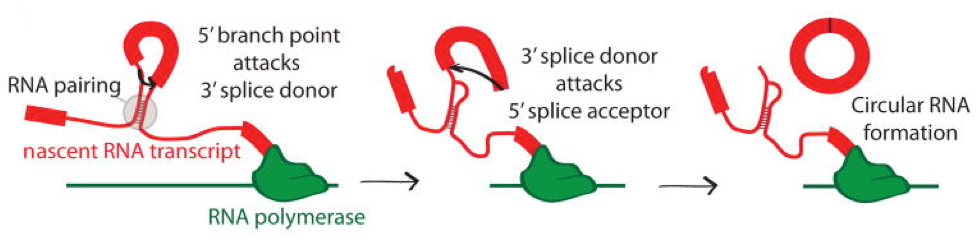 Direct back-splicing: a branch point in the 5’ intron attacks the splice donor of the 3’ intron. The 3’ splice donor then completes the back-splice by attacking the 5’ splice acceptor forming a circRNA. Taken from Jeck & Sharpless Nature Biotechnol (2014).
Direct back-splicing: a branch point in the 5’ intron attacks the splice donor of the 3’ intron. The 3’ splice donor then completes the back-splice by attacking the 5’ splice acceptor forming a circRNA. Taken from Jeck & Sharpless Nature Biotechnol (2014).
This study by Kadener & coworkers involves a plethora of technically complex experimental procedures and associated jargon, from which I’ve extracted what I believe to be some key points to share. After annotating the Drosophila circRNA open reading frames (cORFs), which, by definition,h have the potential for translation, they searched for evidence of their translation utilizing previously published ribosome footprinting (RFP). This led to identification of 37 circRNAs with at least one specific RFP read, referred to as ribo-circRNAs.
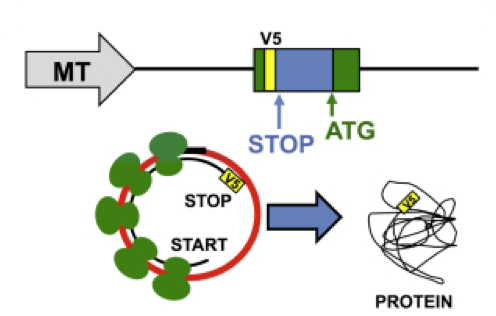 Taken from Jeck & Sharpless Nature Biotechnology (2014)
Taken from Jeck & Sharpless Nature Biotechnology (2014)
Several representative ribo-circRNAs were then constructed to each have (pictured below) a metallothionine (MT) promoter and V5 tag to facilitate translation and anti-V5 antibody-based detection of the expected protein after transfection into cells.
To determine whether circRNAs are translated in a more relevant tissue, they set up the RFP methodology in fly heads. A genetic locus named mbl that is known to produce a circRNA (circMbl3) at high abundance was selected for targeted mass spectrometry from a fly head immunoprecipitated MBL. They utilized synthetic peptides to determine characteristic spectra for which to search in the fly head immunoprecipitate and found a consistent and very high confidence hit for a peptide that can only be produced by circMbl3.
Kadener & coworkers extended these fly head findings to mammalian mouse and rat systems, but the most interesting part of this study—in my opinion—dealt with what signals ribosome binding and translation in the absence of the 5’ cap structure present in mRNA. They demonstrated circRNA translation under conditions intended to block normal 5’ cap-dependent translation of mRNA, and concluded that “[untranslated regions>
of ribo-circRNAs (cUTRs) allow cap-independent translation [and that>
further research is necessary to uncover how these sequences promote translation.”
Remarkably, as you’ll now read, another group of investigators have apparently found how such promotion of circRNA translation can occur.
Translation of circRNA is Driven by N6-Methyladenosine (m6A)
The most abundant modification of RNA in eukaryotes is m6A, which has been recently shown by Li et al. to recruit binding proteins that collectively facilitate the translation of specifically targeted mRNAs—i.e. those “marked” with m6A—through interactions with 40S and 60S ribosome subunit “machinery” that actually carry out translation. Contemporaneously, Yang et al. found that m6A likewise promotes efficient initiation of protein translation from circRNAs in human cells. They discovered that consensus m6A motifs are enriched in circRNAs, and a single m6A site is sufficient to drive translation initiation.
As depicted below, this m6A-driven translation requires initiation factor F4G2 and m6A “reader” YTHDF3. Experiments showed that this translation is enhanced by methyltransferase METTL3/14 and inhibited by demethylase FTO, which enzymatically “add” and “subtract” methyl (Me) groups on specific adenosines (A) in circRNAs, respectively. It has also been shown to be upregulated upon heat shock, which is a commonly employed method to induce “stress” in cells.
Further analyses through polysome profiling, computational prediction and mass spectrometry revealed that m6A-driven translation of circRNAs is widespread, with hundreds of endogenous circRNAs having translation potential. Yang et al. concluded by stating that their “study expands the coding landscape of [the>
human transcriptome, and suggests a role of circRNA-derived proteins in cellular responses to environmental stress.”
Zinc Finger Protein in Muscle Cell Development
Finally, and essentially contemporaneously with above mentioned two publications, a third independent investigation reported by Legnini et al. demonstrated selective circRNA downregulation using short-interfering RNAs (siRNAs). These reagents for RNA interference (RNAi) were used in an image-based functional genetic screen of 25 circRNA species, conserved between mouse and human, expression of which are differentially expressed during myogenesis (i.e. formation of muscular tissue) in Duchenne muscular dystrophy myoblasts.
This siRNA/RNAi-based functional analysis provided one interesting case related to zinc finger protein 609 (circ-ZNF609)—a reported miRNA sponge—the phenotype of which could be specifically attributed to the circular form and not to the linear mRNA counterpart. Consistent with the circ-ZNF609 sequence having an ORF, they found that a fraction of circ-ZNF609 RNA is loaded onto polysomes and that, upon puromycin treatment, it shifted to lighter fractions, similar to mRNAs. The coding ability of this circRNA was proved through use of artificial constructs expressing circular tagged transcripts, and by CRISPR/Cas9—the trendy gene editing method about which I’ve already commented multiple times.
Despite all this evidence, Legnini et al. stated that they “have no hints on the molecular activity of the proteins derived from circ-ZNF609 and as to whether they contribute to modulate or control the activity of the counterpart deriving from the linear mRNA.”
In thinking about closing comments about this update in circRNA, I decided to emphasize that investigations in the field of RNA continue to reveal complexities that will require many more years of global attention to unravel and understand. In just the past decade or so we’ve learned about gene regulation by miRNA/siRNA, reclassification of “junk DNA” as encoding a myriad of long noncoding RNA (lncRNA), mRNA regulation by base-modifications, and curious circRNAs that are more than sponges, and likely encode hundreds (if not thousands) of proteins whose functions have yet to be elucidated. Amazing!
What are your thoughts about all of this?
Your comments are welcomed.
Postscript
After writing this blog, Panda et al. at the National Institute on Aging-Intramural Research Program, National Institutes of Health published a paper titled High-purity circular RNA isolation method (RPAD) reveals vast collection of intronic circRNAs. Here’s a snippet of the abstract which adds to the increasingly curious occurrence of circRNAs that begs, if you will, further research aimed at discovering functions of circRNA-derived proteins.
“Here, we describe a novel method for the isolation of highly pure circRNA populations involving RNase R treatment followed by Polyadenylation and poly(A)+ RNA Depletion (RPAD), which removes linear RNA to near completion. High-throughput sequencing of RNA prepared using RPAD from human cervical carcinoma HeLa cells and mouse C2C12 myoblasts led to two surprising discoveries: (i) many exonic circRNA (EcircRNA) isoforms share an identical backsplice sequence but have different body sizes and sequences, and (ii) thousands of novel intronic circular RNAs (IcircRNAs) are expressed in cells. In sum, isolating high-purity circRNAs using the RPAD method can enable quantitative and qualitative analyses of circRNA types and sequence composition, paving the way for the elucidation of circRNA functions.”





{kind=link}
{kind=link}
{kind=link}